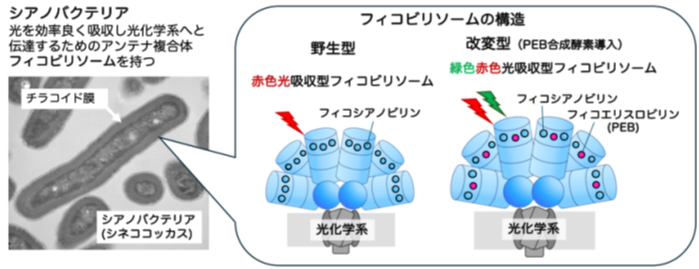
埼玉大学,理化学研究所,東京大学は,光合成微生物であるシアノバクテリアを用いて,糖脂質の前駆体であるジアシルグリセロール(DAG)をリン脂質の前駆体であるホスファチジン酸(PA)へ変換する DAGキナーゼ(DGK)の変異株を作出し,強光ストレス応答における膜脂質転換の役割について解明した(ニュースリリース)。
光合成生物は,環境ストレスに応答することで,光合成活性を最適化している。光合成の場であるチラコイド膜においては,強光や低CO2などの環境変化に応じて膜脂質組成が変化することで光合成活性が制御されていると考えられるが,その分子機構については不明である。
シアノバクテリアの膜脂質合成経路においては,リン脂質の前駆体であるホスファチジン酸(PA)と糖脂質の前駆体であるDAGが,DGKとPAホスファターゼ(PAP)によって相互に変換されることで,リン脂質と糖脂質の割合を調整していると考えられる。
そこで,大腸菌のホモログ探索から,シアノバクテリアSynechocystis sp. PCC 6803においてDGKをコードすると考えられる遺伝子の探索を行ない,遺伝子を耐性遺伝子の挿入によって破壊した株を作出した。
遺伝子破壊株では,DAGからPAの変換が行なわれなかったことから,Synechocystisにおいて,DGKをコードする遺伝子を同定することができた。野生株においては,強光下において,PG量が約1.7倍程度に増加することが観察されたが,DGK遺伝子を破壊したdgkA株では,強光下におけるPGの増加が見られず,MGDGが2倍に増加した。
また,dgkA変異株は強光下で生育遅延を示した。そこで,強光下における光合成活性を解析したところ,dgkA変異株では,水分解反応を担う光化学系II(PSII)の修復が阻害されていた。また,dgkA変異株では野生株に比べて,PSII二量体が増加しており,PSIIの修復過程における複合体の解体過程が阻害されている可能性が見出された。
また,これらの表現型は,細胞懸濁液にPGを添加することによって,改善されたことから,dgkA変異株ではMGDG/PG比の減少によって,PSII修復阻害が引き起こされたと考えられる。
研究グループは,今後は,強光以外の環境ストレスに関わる膜脂質転換メカニズムを明らかにすることによって,強光だけでなく,さまざまな環境ストレスに対して耐性を持つ光合成系の構築に展開したいとしている。