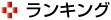 神戸大学と茨城大学,神奈川大学を中心とした研究グループは,近赤外光を有効活用する光合成細菌のキメラ複合体発現系の構築およびその特性解析を行ない,エネルギー勾配を遡って低い光エネルギーを高いエネルギーに変換する分子機構の一端を明らかにした(ニュースリリース)。
神戸大学と茨城大学,神奈川大学を中心とした研究グループは,近赤外光を有効活用する光合成細菌のキメラ複合体発現系の構築およびその特性解析を行ない,エネルギー勾配を遡って低い光エネルギーを高いエネルギーに変換する分子機構の一端を明らかにした(ニュースリリース)。
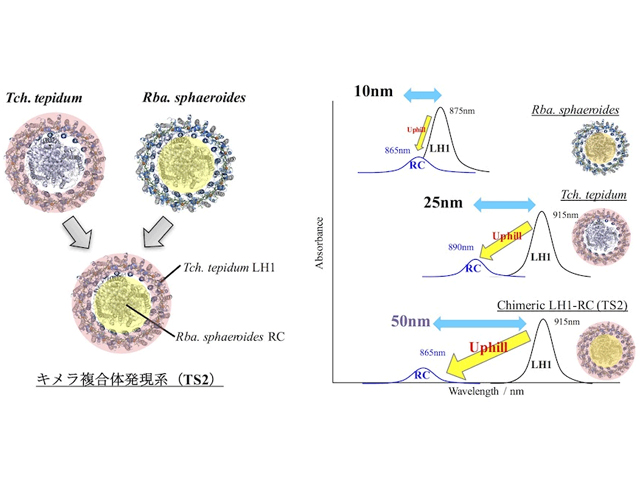
紅色光合成細菌「サーモクロマチウム・テピダム (Thermochromatium tepidum)」(Tch. tepidum)は,近赤外光を利用して光合成を行なうことができる紅色光合成細菌の中でも,非常に低エネルギーの近赤外光を吸収することが可能で,エネルギーの勾配に逆らうuphill型のエネルギー移動を行なっている。
近年の研究から,Tch. tepidumのLH1-RC複合体(紅色細菌が持つ光合成を行なうための装置で,光を集める集光性アンテナタンパク質(LH1)と光合成反応を行なう反応中心(RC)の複合体)は,LH1のC末端領域にCa2+(カルシウムイオン)の結合サイトを有しており、Ca2+と色素タンパク質との相互作用によって低エネルギー吸着特性を示すことがわかってきた。しかし分子機構など詳細については明らかにされていない。
研究では,このLH1がTch.tepidum由来,RCがRba.sphaeroides由来であるキメラ複合体発現系(TS2)およびCa2+配位子と推定されるTch.tepidum LH1のアミノ酸(α-Asp49,α-Asn50,α-Leu46)に部位特異的変異を導入した各種変異体について分光学的解析を行ない,Tch.tepidum,Rba.sphaeroidesとの比較研究を行なうことで,Ca2+結合サイトとLH1の低エネルギー吸収特性との関連性,およびuphill型エネルギー移動の分子機構を解明することを目的とした。
はじめに,TS2および部位特異的変異体由来光合成膜小胞について,吸光分光法,近赤外および可視共鳴ラマン分光法により,色素分子の吸収特性や,LH1タンパク質とBChl-a分子およびカロテノイド分子間の相互作用を解析した。
全ての変異株は嫌気条件で光合成生育可能であり,変異に応じて吸収特性が顕著に変化した。TS2ではTch.tepidumと同じBChl-aの吸収バンドを示したが,カロテノイドはRba.sphaeroidesと同じスフェロイデンのバンドが観測されたことから,TS2のLH1ではBChl-aはTch.tepidumと同じだが,カロテノイドがスピリロキサンチンからスフェロイデンに変化していることがわかった。
また,この変化によってTS2におけるカロテノイドからBChl-aへのエネルギー移動効率はTch.tepidumよりも高くなっており,光合成効率を向上させる効果があることが判明した。
近赤外共鳴ラマン分光法による解析の結果,TS2の異常な低エネルギー吸収特性はTch.tepidum同様,BChl-aのカルボニル基とLH1タンパク質のTrpの間に働く水素結合の強さに依存していることが明らかになった。一方,Tch.tepidumのLH1 Ca2+結合サイトは,α-Asp49,α-Asn50,α-Trp46,β-Leu46で構成されると考えられていたが,α-Asn50がCa2+の配位子でない可能性が高いことを強く支持するデータが得られた。
この結果は最近報告された高分解能の結晶構造によって証明された。また,α-43位の欠損がCa2+結合サイトの形成において極めて重要であることが判明した。