 横浜市立大学と米国国立衛生研究所は,転写因子IRF8による貪食細胞への分化決定の分子メカニズムを解明した(ニュースリリース)。
横浜市立大学と米国国立衛生研究所は,転写因子IRF8による貪食細胞への分化決定の分子メカニズムを解明した(ニュースリリース)。
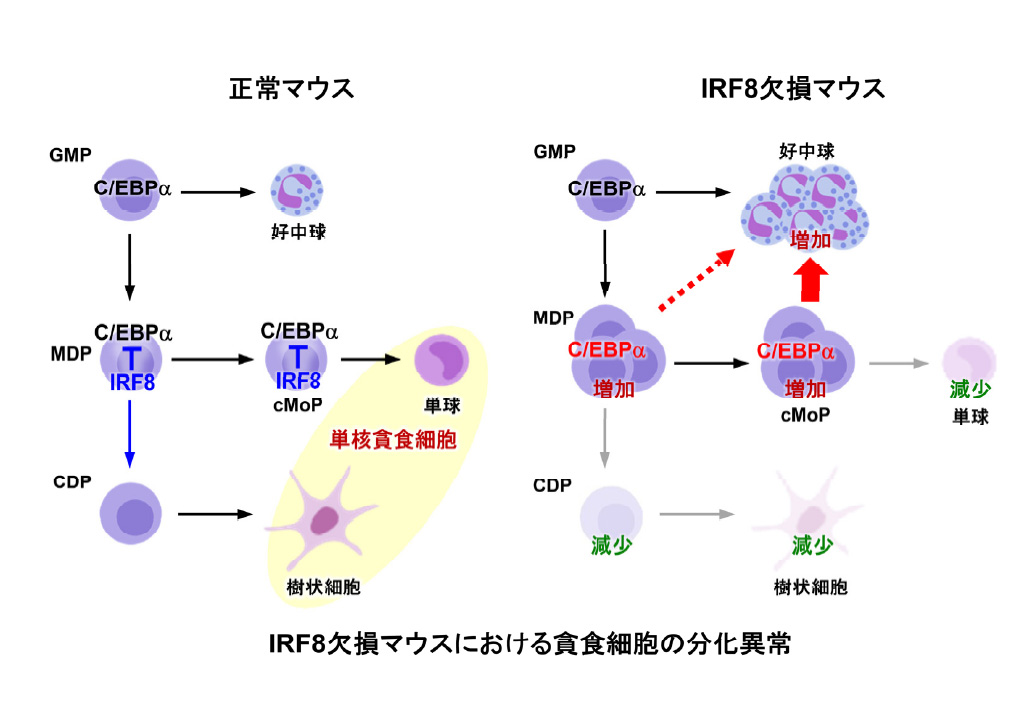
白血球のうち単核貪食細胞は,感染の際に病原微生物を貪食する細胞で,その微生物の情報を獲得免疫系のT細胞やB細胞に伝える能力を持っている。単核貪食細胞が産生されるためには,造血幹細胞から骨髄系共通前駆細胞,顆粒球-単球前駆細胞(GMP),単球-樹状細胞前駆細胞(MDP)という段階を経ることが知られている。
MDPの下流には単球前駆細胞(cMoP)と樹状細胞共通前駆細胞(CDP)が存在し,それぞれ単球と樹状細胞を産生する。ここでGMPがMDPになるためには顆粒球(好中球など)への分化能を失う必要があるが,その仕組みは不明だった。
転写因子IRF8の遺伝子変異をもつヒトや遺伝子を欠損したマウスでは単球や樹状細胞が著しく減少する一方で好中球が異常に増加する。実際IRF8は好中球分化を抑制することが示されている。しかしながら,IRF8が具体的にどの前駆細胞でどのようなメカニズムで好中球分化を抑制するのかはわかっていなかった。
研究グループはまず造血幹細胞や各種前駆細胞を詳しく調べ,正常なマウスではIRF8タンパク質がMDPの段階から初めて強く発現することを見出した。IRF8欠損マウスではMDPやcMoPが野生型のマウスよりも著しく増加していた。そしてIRF8欠損マウスのMDPやcMoPは樹状細胞や単球の産生能が著しく低下しており,本来なるはずのない好中球へと分化してしまうことを突き止めた。
これは,IRF8欠損マウスでは蓄積したMDPやcMoPから好中球が産生されてしまうことが,大きな原因であると考えられる。その原因を詳細に検討するために,マイクロアレイによる網羅的遺伝子発現解析とバイオインフォマティクスによるパスウェイ解析を行なったところ,IRF8遺伝子欠損マウスのMDPやcMoPの細胞内部ではC/EBPαという別の転写因子が異常に活性化していることがわかった。
C/EBPαはヒトやマウスの顆粒球の分化・産生に必須の転写因子。そこでIRF8とC/EBPαを細胞に発現させたところ,IRF8はC/EBPαによる転写活性化や好中球への分化誘導を強く抑制することがわかった。さらにIRF8はMDPやcMoPにおいてC/EBPαに直接結合し,C/EBPαが好中球への分化に重要な標的遺伝子のDNAに結合するのを防ぐことがわかった。
最後にIRF8欠損マウスの前駆細胞にC/EBPαの機能を弱める変異型C/EBPを発現させたところ,IRF8欠損マウスに認められた好中球の異常産生が正常マウスのレベルまで戻ることがわかった。すなわちIRF8はC/EBPαの機能抑制によって,単核貪食細胞前駆細胞が好中球にならないようにしていることがわかった。
研究グループでは今回の研究について,細胞分化の基本的な仕組みを理解する上でも大切な知見であり,他の細胞種においても類似の仕組みが用いられている可能性があるとしている。また慢性骨髄性白血病やある種の原発性免疫不全症ではIRF8の発現や機能の低下が起こり,樹状細胞の分化・産生が低下する一方で好中球が著増することが知られており,今回の発見はこれらの疾患の発症機序や病態の理解,そして新たな治療法を開発するためにも重要な知見であるとしている。
関連記事「東工大ら, 四肢形成時に細胞の生死の運命を決めるメカニズムを解明」「東大ら,細胞に「食べる」機能を付与することに成功」「九大,神経障害性疼痛の原因となるタンパク質を同定」